The early history of microbiology
by Anders Skovly, 2026
Introduction
In this article we will look at four different topics: the discovery of microbial multiplication by division, the century-long debate on spontaneous generation, the role of yeast in alcoholic fermentation, and the role of bacteria in infectious disease, the latter in conjunction with the discovery of solid culture methods.
The discovery of multiplication by division
The ability of microorganisms to multiply by dividing themselves in two was first seen by Abraham Trembley, who described two different examples in a letter he sent to the Royal society in 1744 (available at biodiversitylibrary.org). The first example was from an organism Trembley called the “clustering Polypus”. In his 1744 letter he introduces this organism as follows:
We find, in divers places, upon water-plants, and other bodies in the water, a whitish substance, that looks at first only like a sort of mould: We sometimes see plants, sticks of wood, snail-shells, and the like, that are entirely covered over with this substance. But if we take any of these, put them into a glass of clear water, and then examine with a magnifying glass what is upon them, we soon discover, in the little bodies, that, by their assemblage, form this whitish substance, such motions as give sufficient reason to look upon them as living animals; and this will appear yet more sensible, when they come to be observed with a microscope. We then find them to be minute bodies, severally fixed to the extremities of small stems, or pedicles, many of which are often so united, as to form together a sort of branches, or clusters;
Here is a modern micrograph of clustering polypi growing on a filamentous green alga.
Further on he gives a detailed description of how the clustering Polypi multiply:
These Polypi are capable of swimming about; and when they swim, they are no longer in clusters, but always single; and they do not then appear in the same form as when they were fixed, and open at their anterior ends. It is by swimming that they leave the place to which they first appeared fixed, and that they go and fix themselves to any other body that they find in their way.
One should begin to observe a Polypus soon after it has fixed itself singly, in order to see regularly in what manner the clusters form themselves, and in what way these small creatures multiply.
The stem or pedicle of a Polypus that is yet single, and which has but lately fixed itself, is at first very short, but it lengthens itself in a little time. After that, the Polypus multiplies; that is to say, it divides or splits itself into two lengthwise. One first observes the lips to be drawn into the body, whose anterior part closes, and becomes round: the motion that was to be seen before the lips were drawn in, no longer appears; yet one may see, by looking with attention, a slow motion within the body, during all the time that the Polypus remains closed. The anterior part of the Polypus flats itself afterwards by degrees, and spreads in proportion, becoming broader as it shortens; it then gradually splits down through the middle, that is, from the middle of the head to the place where the posterior end joins to the pedicle: so that, in a little while, there appear two separate round bodies joined to the extremity of the pedicle that just before supported but one.
The anterior part of each of these bodies then opens by degrees; and, as they open, the lips of the new Polypi shew themselves more and more. Then is the time of observing these lips with attention, and of forming to one's self an idea of their true form, and of their motion already spoken of. This motion is at the first very slow, it quickens as the Polypi continue to open; and, as soon as they have done, it becomes as swift as that which appeared in the lips of the whole single Polypus, before it began to divide: and then these new Polypi may be looked upon as entirely formed. They are, at first, less than the Polypus from which they were formed; but they grow to the same size in a very little time.
A Polypus is an hour, or thereabout, dividing itself. To have a tolerable notion of this operation, one must have seen it divers times, and in Polypi placed and situated in divers different ways.
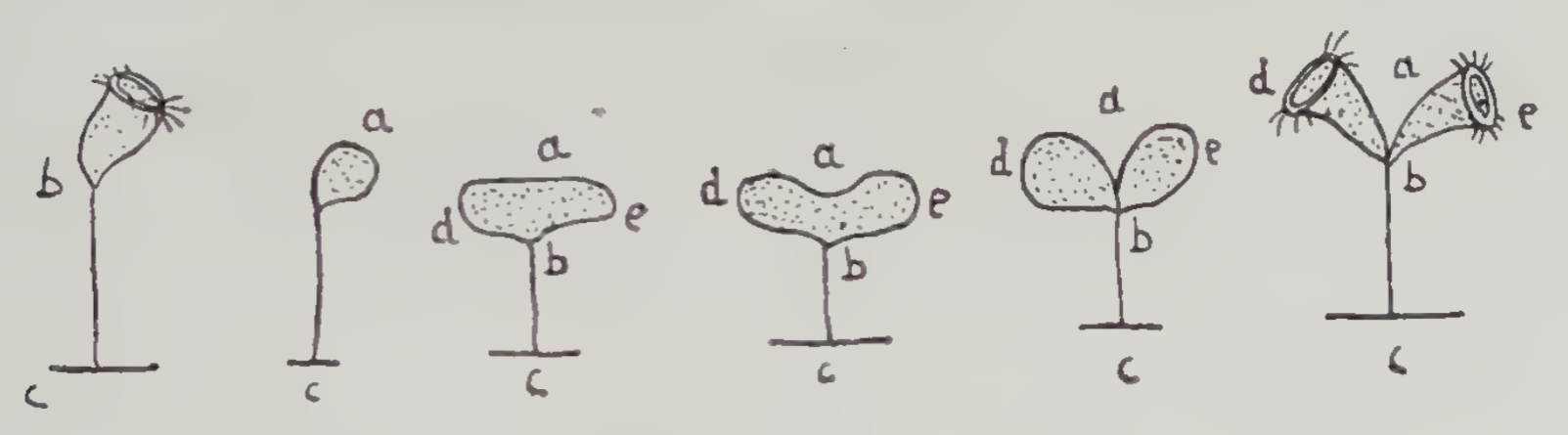
Trembley did not include this drawing in the mentioned 1744 letter. Instead, my source for this drawing is John Baker's biography Abraham Trembley of Geneva: Scientist and Philosopher (1952). Baker gives as his source the book Correspondance inédite entre Réaumur et Abraham Trembley comprenant 113 lettres recuetllies et annotées by Maurice Trembley (1943).
(According to John Baker, the modern name of the “clustering Polypus” is Epistylis anastatica.)
In his 1744 letter Trembley also describes division of a second organism, which he called a “funnel-like Polypus”. This creature divides in a sort of diagonal manner.
Here is a modern micrograph of funnel-like Polypi.
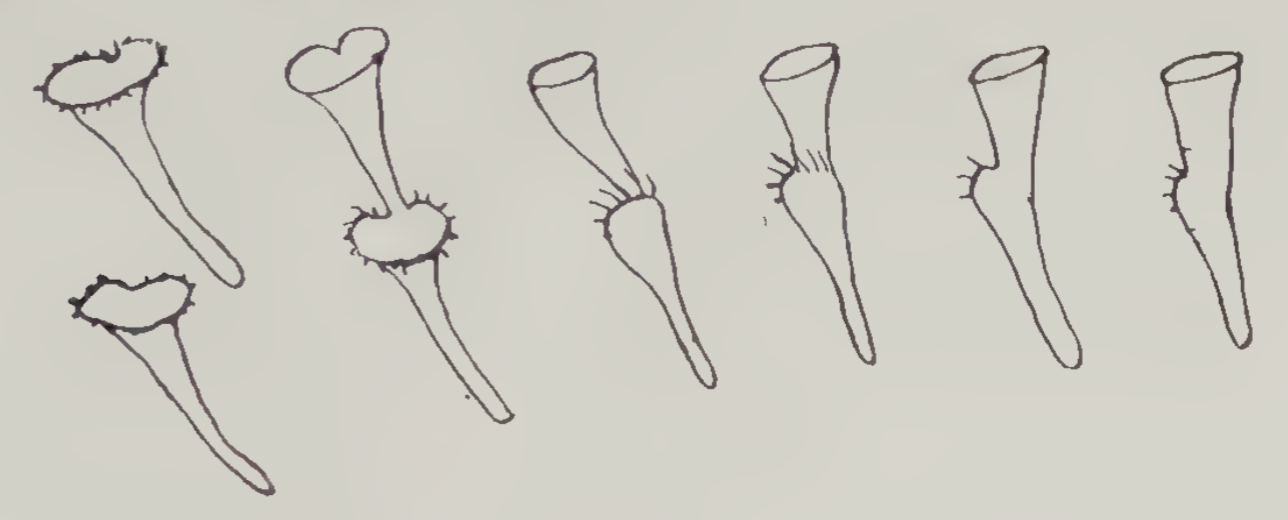
Source: see the previous figure.
(According to John Baker, the modern name of the “funnel-like Polypus” is Stentor.)
Spontaneous generation
If you put some beans into water, boil the water, and let it cool off, then all sorts of microorganisms will develop in the water. The same occurs if instead of the beans you use another vegetable, or some meat.
These microorganisms could not be seen in the water prior to the boiling. And if these organisms developed from some kind of sub-microscopic egg or seed, then presumably these eggs or seeds should have been destroyed by the boiling of the water. So, where does these microscopic beings come from?
Some people believed the answer was “spontaneous generation”, that is, the creation of living organisms from non-living matter. For example, if you leave beans in water, then some of the bean matter will dissolve into the water. One can hypothesize that the sub-microscopic bean particles in the water have a life-force, which allow them to assemble into living organisms. This is the hypothesis of spontaneous generation.
Spontaneous generation was a point of contention among biologists for well over a century, until it was finally disproved in 1862 by Louis Pasteur. His work on the topic was published in an article titled “Memoir on the organized bodies which exists in the atmosphere. Examination of the doctrine of spontaneous generation.” (“Memoire sur les corpuscules organisés qui existent dans l'atmosphère. Examen de la doctrine des générations spontanées.”). The french original is available at archive.org. I have used Google translate to create an English translation, available here.
Let's take a look at Pasteur's memoir. The first chapter gives some historical background on spontaneous generation:
In antiquity and until the end of the Middle Ages, everyone believed in the existence of spontaneous generation. Aristotle said that any dry body that becomes wet and any wet body that dries out will give rise to animals [...] Such errors could not long withstand the spirit of inquiry that gripped Europe in the 16th and 17th centuries. Redi, a famous member of the Accademia del Cimento, demonstrated that the worms in rotting flesh were fly larvae. His proofs were as simple as they were decisive, for he showed that simply wrapping rotting flesh in fine gauze was enough to absolutely prevent the birth of these larvae. Redi was also the first to recognize males, females, and eggs in the animals that live in other animals. These flies that lay their eggs in fruit were caught in the act, Réaumur later said, and it was understood, when a worm was seen in an apple, that it was not the rot that had engendered it, but rather that the worm was the cause of the fruit's decay.
But soon, in the second half of the 17th century and the first half of the 18th, microscopic observations proliferated. The doctrine of spontaneous generation then reappeared. Unable to explain the origin of these diverse organisms revealed by the microscope in infusions of plant or animal matter, and seeing nothing resembling sexual generation in them, some were led to believe that matter which had once lived retained its own vitality after death, under whose influence its disjointed parts reunited, under certain favorable conditions, with variations in structure and organization determined by those very conditions. Others, on the contrary, adding to the marvelous results revealed by the microscope through their imagination, believed they saw mating in these infusoria* — males, females, eggs — and positioned themselves as declared opponents of spontaneous generation. It must be admitted that the evidence supporting either of these opinions hardly stood up to scrutiny.
* “Infusoria” refers to those microscopic organisms which typically develops in meat infusions and plant infusions. As with “animalcules”, the term “infusoria” is not in use in modern biology.
The question stood at this point when, in 1745, a work by Needham appeared in London. Needham was a skilled observer and a devout Catholic priest, a circumstance which, in such a subject, seemed to guarantee the sincerity of his convictions. In this work, the doctrine of spontaneous generation was based on facts of an entirely new kind: experiments on hermetically sealed vessels, previously exposed to the effects of temperature. It was Needham, in fact, who first conceived of such experiments.
At this point, let us leave Pasteur for a moment, and take a look at Needham's experiment.
Needham’s experiment
John Needham's work was published in a 1749 book titled “Observations upon the generation, composition, and decomposition of animal and vegetable substances”. This book is available on biodiversitylibrary.org. (I guess this work had also been published in 1745, as Pasteur said, but I only found this 1749 version.)
Needham writes that many people at the time believed animalcules (microorganisms) to originate from eggs:
It has generally been thought by naturalists, that microscopical animalcules were generated from eggs transported through the air, or deposited by a parent fly, invisible to the naked eye, or even that assisted with microscopes. Yet is it strange that no naturalist should yet have seen them, if they are really so numerous, when their supposed progeny is so various, and themselves must be thought to be so frequently gliding over the surface of all stagnant waters.
To test whether animalcules can originate from eggs, Needham performs an experiment where he fills a vial with a meat infusion, seals the vial with a cork to prevent contamination from the surrounding air, and heats the vial to destroy any hypothetical animalcule-eggs in the infusion. He then leaves the vial alone for some time, before opening it to check with a microscope whether any animalcules have developed inside. Here’s how Needham describes his experiment:
For my purpose therefore, I took a quantity of mutton-gravy hot from the fire, and shut it up in a phial, clos'd up with a cork so well masticated, that my precautions amounted to as much as if I had sealed my phial hermetically. I thus effectually excluded the exterior air, that it might not be said my moving bodies drew their origin from insects, or eggs floating in the atmosphere. I would not instil any water, lest, without giving it as intense a degree of heat, it might be thought these productions were convey'd through that element [...] I neglected no precaution, even as far as to heat violently in hot ashes the body of the phial; that if any thing existed, even in that little portion of air which filled up the neck, it might be destroy'd, and lose its productive faculty. Nothing therefore could answer my purpose of excluding every objection, better than hot roast-meat gravy secur'd in this manner, and exposed for some days to the summer-heat: and as I was determined not to open it, till I might reasonably conclude, whether, by its own principles, it was productive of any thing, I allowed sufficient time for that purpose to this pure unmix'd quintessence, if I may so call it, of an animal body.
... My phial swarm'd with life, and microscopical animals of most dimensions, from some of the largest I had ever seen, to some of the least. The very first drop I used, upon opening it, yielded me multitudes perfectly form'd, animated, and spontaneous in all their motions ...
I shall not at this present time trouble you with a detail of observarions upon three or four scores of different lnfusions of animal and vegetable substances, posterior to these upon mutton-gravy; all which constantly gave me the same phenomena with little variation, and were uniform in their general result ...
So, where does Needham think the animalcules in his vials come from, if not from som kind of eggs? His hypothesis is that organic particles contain a “vegetative force” which allows these particles to assemble into microscopic organisms:
... It seems plain therefore, that there is a vegetative force in every microscopical point of matter, and every visible filament of which the whole animal or vegetable texture consists: and probably this force extends much farther; for not only in all my observations, the whole substance, after a certain separation of salts and volatile parts, divided into filaments, and vegetated into numberless zoophytes, which yielded all the several species of common microscopical animals; but these very animals also, after a certain time, subsided to the bottom, became motionless, resolv'd again into a gelatinous filamentous substance, and gave zoophytes and animals of a lesser species.
Needham versus Spallanzani
We can now return to where we left off in Pasteur's “Memoir on the organized bodies which exists in the atmosphere”:
... But Needham's conclusions were soon subjected to experimental verification. At that time, there was in Italy one of the most skilled physiologists science could boast, the most ingenious, the most difficult to please: Abbé [Lazzaro] Spallanzani.
It would be pointless to present a complete history of the quarrel between the two naturalists. But it is important to clearly define the experimental difficulty to which they particularly applied their efforts, and to determine whether this long debate had dispelled all doubts. This is generally believed. Spallanzani is readily regarded as Needham’s victorious adversary. An impartial examination of their contradictory observations on the most delicate point of the subject will show us that Needham could not, in all fairness, abandon his doctrine in the face of Spallanzani’s work.
“It only remains for me,” said Needham, “to speak of Spallanzani's latest experiment, which he himself considers the only one in his entire dissertation that seems to have any force against my principles.”
“He [Spallanzani] hermetically sealed nineteen vessels filled with various plant substances and boiled them, thus sealed, for an hour. But from the way he treated and subjected his nineteen plant infusions to such rigorous testing, it is clear that he not only greatly weakened, or perhaps completely destroyed, the vegetative force of the infused substances, but also that he entirely corrupted, through the exhalations and the heat of the fire, the small portion of air that remained in the empty part of his flasks.”
So these are Needham’s objections: first, that the lack of animalcules in Spallanzani’s plant infusions may be due to the destruction of the hypothetical “vegetative force” by prolonged boiling (1 hour).
And second, that the lack of animalcules may be due to the poor air quality in the flasks. While Needham sealed his vials with cork, Spallanzani used a flame to melt the glass in the neck of his flasks, thus creating a hermetic glass seal. Needham considers the possibility that Spallanzani’s flame has somehow damaged the air in the flasks, or that the flame has heated the air to such extent that the air was expelled from the flasks during the sealing.
Spallanzani's experiments
Spallanzani responded to Needham's objections with a new series of experiments, and these are rather interesting. The experiments are described in chapters 1 and 2 of Spallanzani's 1776 book “Tracts on the natural history of animals and vegetables, Volume 1“ (“Opuscoli di fisica animale, e vegetabile, Volume Primo”). An english translation by John Dalyell is available on biodiversitylibrary.org.
First, to investigate if prolonged boiling destroys the “vegetative power” of infusions, Spallanzani creates 32 plant infusions. 8 infusions are boiled for half an hour, 8 are boiled for an hour, 8 are boiled for a hour and a half, and 8 are boiled for two hours. These vessels were not hermetically sealed, but were “loosely stopped with corks”. Here is his description of the results:
On the 15 of September, I made thirty-two infusions; and on the 23 examined them for the first time. Animalcula were in all; but the number and species different in each. In the maize infusions, they were smaller, and proportionally more rare, according to the duration of boiling.
... In the infusion of kidney beans, boiled two hours, were three species of animalcula; very large; middle sized; and very small. The figure of the first, partly umbellated and attached to long filaments dragged along in their progress; the second were cylindrical; and the third, globular. All three were incredibly numerous. In the infusion boiled [a hour an a half], were animalcula of the largest and smallest class, but few in number; still fewer, in that boiled an hour; and fewest of all, in that boiled half an hour.
(Then he continues to describe the appearances of the animalcules in the other six infusions.)
... Thus, it is clearly evident, that long boiling of seed infusions does not prevent the production of animalcula; and, notwithstanding the maize does not seem to favour it, four infusions strongly corroborate the fact.
After the boiling experiments Spallanzani wanted to test infusions created from seeds exposed to higher temperatures than that of boiling water. He thus creates infusions from seeds heated in a coffee roaster, and found that such infusions also gave rise to animalcules. He continues:
Vegetable seeds were exposed to trials more severe: they were exposed to the greatest heat that can be excited by common fires, or fire augmented by art: Burning coals, and the flame of the blow pipe, were the two agents exercising their power on them. And, in the first place, I kept them on an iron plate above burning coals until entirely consumed by the violence of the flames, and converted to a dry cinder, which was reduced to powder, and as many infusions formed as there were seeds. A cinder was also made by the blow pipe, which, besides excessive aridity, had acquired considerable hardness. I must acknowledge I did not in the least expect to find animalcula in this new infusion. After viewing them once and again, hardly able to credit my eyes, I repeated the experiment twice. Some suspicion arose that the animalcula might come from the water used rather than the burnt seeds; therefore, on repeating the experiment, the same as what formed the infusions was put in other vessels. Both times, however, they re-appeared in the burnt seeds, while not one was seen in the water.
These facts fully convinced me, that vegetable seeds never fail to produce animalcula, though exposed to any degree of heat; whence arises a direct conclusion, that the vegetative power is nothing but the work of imagination; and if no animalcula appear in vessels hermetically sealed and kept an hour in boiling water, their absence must proceed from some other cause.
In chapter 2 of his book, Spallanzani describes further experiments with infusions contained in hermetically sealed vessels. Remember that Spallanzani had sealed his flasks by using a flame to melt the glass neck, producing a solid glass seal. Needham had objected that the flame may cause damage to the air inside the flasks. To try to counter this objection, Spallanzani produced flasks with very thin necks, which could be sealed by the flame very quickly:
... the neck of the vessel was drawn out at the [flame] almost to a capillary tube: the smallest part was then instantaneously sealed, so that the internal air could suffer no alteration, as was evident from no hissing being heard on breaking the vessel ...
A set of nine seed infusions were thus hermetically sealed in flasks, and the flasks were placed in boiling water for half a minute. Another set of nine hermetically sealed infusions were boiled for a minute, a third set boiled a minute and a half, and a fourth set boiled two minutes. That’s 36 flasks total.
For comparison, another 36 seed infusion flasks were boiled in the same way, except that these other flasks were not hermetically sealed. After eleven days the non-sealed flasks were full of animalcules. Spallanzani then broke the sealed flasks and examined their contents, to see whether they too had developed animalcules.
Of the nine types of seed infusions which had been hermetically sealed and boiled, three infusions produced no animalcules (not for any of the boiling durations.) The other six types of infusions, which had been made from beans, vetches, buck-wheat, mallows, maize, and lentils, all contained animalcules (for all four boiling durations). But these animalcules were all of the extremely small kind, and none of the relatively larger animalcules were present.
From this series of experiments I concluded, that the heat of boiling water half a minute was fatal to large, middle sized, and small animalcula [...] while the same heat, protracted two minutes, did not affect the infinitely minute animalcula ...”
(Thomas Brock, in his book “Milestones in Microbiology”, considers that the “infinitely minute animalcula” were bacteria, while the “large, middle sized, and small animalcula” were protists.)
Spallanzani then goes on to test what boiling time is required to completely prevent the development of the very small animalcules. He finds that half an hour of boiling is not enough, while a little less that three quarters of an hour is enough.
He also finds that the development of “large, middle sized, and small animalcules” can be prevented by placing the infusion flasks in boiling water for just a single second. On further examination he discovers that if the hermetically sealed infusion flasks are placed in water which is slowly heated to 95°F (35°C), after which the flasks are removed from the water and stored for a while, the “large, middle sized, and small animalcules” do not develop. But when the water is only heated to 93°F (33.9°C), these animalcules do develop.
Based on all the results, he thinks his experiments disproves the idea of a “vegetative force” as the source of creation of animalcules.
Thus we are led to ascribe their origin to included germs, which for a limited time can resist the influence of heat, but at length yield under it. As the [large, middle sized, and small animalcules] does not originate unless at a much more moderate degree of heat, there is a necessary inference, that the germs of [these animalcules] are much sooner affected than those of [the extremely small animalcules]; whence we must conclude, that the amazing multitude of animalcula in the infusions of open vessels exposed to boiling heat, and the intense flame of the blow pipe, do not appear because their germs have resisted so great a degree of heat, but because other germs have come to the infusions after cessation of the heat.
But is there any proof, or any forcible argument, to remove or lessen our natural repugnance to suppose that germs of the [extremely small] animalcula can resist the heat of boiling water? To speak of the germs or eggs of animals known to us, are there none of this nature? Undoubtedly, as far as our knowledge extends, we are unacquainted with any of that description. Something has been said on the subject in the ninth chapter of my Dissertation, demonstrating how the eggs of various insects, as well as those of birds, perish at a degree of heat considerably less than that of boiling water. It is further shown, that this heat injures the seeds of plants, even those with the hardest shell. A greater number of seeds and eggs, indeed, might have been subjected to experiment: and some may be found capable of standing the trial. With regard to seeds, hope should not be abandoned from reading in Duhamel, that he succeeded in the germination of wheat which had suffered [234°F (112°C)] in a stove; and it is very probable this seed is not unique.
So we see, that Spallanzani hypothesizes that the very small animalcules develop from some kind of eggs capable of withstanding the heat of boiling water for a long time. However, he's not able to add any kind of evidence or support to this hypothesis. But he was certainly on to something: a century later, other scientists would discover that some bacteria can create and release spores. These spores are very resistant to heat, and can later germinate and develop back into bacteria.
Some further developments
Again we return to Pasteur's “Memoir on the organized bodies which exists in the atmosphere”:
... This, certainly, is the clearly defined debate between [Needham and Spallanzani]. It is in chapter 1 [and 2] of the first volume of his Opuscules that Spallanzani addresses this decisive difficulty. And what is his conclusion? To eliminate the production of infusoria [microorganisms which develop in infusions], it is necessary to maintain the infusions at boiling water temperature for three-quarters of an hour.
However, didn't this required duration of a temperature of 100°C for three-quarters of an hour justify Needham's fears about a possible alteration of the air in the vessels? At the very least, Spallanzani should have included an analysis of this air in his experiments, but science was not yet sufficiently advanced. Eudiometry had not yet been developed. The composition of air was barely understood.
... [Nicolas] Appert applied the results of Spallanzani's experiments [...] to domestic economy [commercial production of hermetically preserved food]. For example, one of the Italian scientist's experiments consisted of placing peas and water in a glass vessel, which was then hermetically sealed, and subsequently kept in boiling water for three-quarters of an hour. This is indeed Appert's method. Now, Gay-Lussac, wishing to ascertain the validity of this method, subjected it to various tests and arrived at the following result:
“One can be convinced,” said Gay-Lussac, “by analyzing the air in bottles in which substances (beef, mutton, fish, mushrooms, grape must) have been properly preserved, that it no longer contains oxygen, and that the absence of this gas is consequently a necessary condition for the preservation of animal and vegetable substances.”
(This occured sometime in the early 1800s.)
Let us consider only the first part of Gay-Lussac’s assertion, namely, that there is no longer any oxygen in Appert’s preserves. Do you not see that this justifies Needham’s fears about an alteration of the air in the vessels in Spallanzani’s experiments? As I said earlier, consequently, Spallanzani had not overcome Needham’s objections.
But an experiment by Schwann brought about a very significant advance in the matter ...
(I read somewhere, I don't remember where (though it may well have been in Pasteur 1862), that the reason why there was no oxygen in Appert's preserved food containers was that the containers included only a small amount of air, so that when the containers were boiled for a long time, all the oxygen in that small amount of air would react chemically with the organic matter of the food.)
Schwann's experiments
In 1837, Theodor Schwann published his “Preliminary report concerning experiments on wine-fermentation and putrefaction” (“Vorläufige Mittheilung, betreffend Versuche über die Weingährung und Fäulnis”). Here he made an interesting contribution to the role of air/oxygen in the development of microorganisms in boiled liquids.
(This is the same Schwann as the one mentioned in my previous article on the history of cell biology, he was the guy who wrote about how the “elementary particles” of animals develop in a similar fashion as the cells of plants.)
(While I found Schwann's original article and attempted to Google-translate it, this proved unfeasible, as it wasn't possible to copy the source text into the translator without a lot of errors, to such degree that the pasted text was utter nonsense. Therefore I rely on a partial english translation by Thomas Brock, published in his book “Milestones in Microbiology”. I would like to mention that I discovered a lot of my sources for this article through Brock's book, and if you would like to learn more about the history of microbiology, “Milestones in Microbiology” is definitely worth checking out.)
In his first experiment, Schwann placed a piece of meat inside a flask partially filled with water. The flask was sealed with a lid, with two glass tubes going through the lid. Outside the flask, a part of each glass tube was submerged in a pool of molten metal, which had a temperature close to the boiling point of mercury (i.e. close to 357°C).
In his first experiment, Schwann placed a piece of meat inside a flask partially filled with water. The flask was sealed with a lid, with two glass tubes going through the lid. Outside the flask, a part of each glass tube was submerged in a pool of molten metal, which had a temperature close to the boiling point of mercury (i.e. close to 357°C).
The water in the flask was boiled, and was allowed to cool. Then, an air pump was used to move air through one of the glass tubes, into the flask, and out the other glass tube. As the two tubes were partially submerged in molten metal, the air was “pre-heated” before it entered the flask. (I don’t think it is stricktly necessary to heat the air that is leaving the flask through the second tube. Heating the air in this exit-tube was perhaps just a precaution to prevent unheated air from entering the flask.) The heated air was cooled down again before actually entering the flask, so this pumping of air did not heat up the water and meat in the flask.
Normally, when water with a piece of meat is boiled and cooled and exposed to air, microorganisms will develop in the water. Schwann found that with his setup, where the air was heated before entering the flask, microorganisms did not develop:
This experiment was performed many times, and in all cases there was no putrefaction or production of infusoria or mold, even after many weeks, and the liquid remained as clear as it had been right after preparation
(That “the liquid remained clear” means that there is no significant development of microorganisms in the liquid, as a liquid full of microorganisms will be “cloudy” and semi-opaque.)
... I will only remark here, that this experiment, when viewed from the standpoint of an opponent of spontaneous generation, can be explained as follows: the germs of molds and infusoria, which according to [the alternative hypothesis] are present in the air, are killed by the thorough heating of the air.
In a second experiment, four flasks were filled with water, sugar, and beer yeast. The flasks were sealed and placed in boiling water until the water inside the flasks also began to boil. The flasks were then removed from the the boiling water, and were placed upside down in a pool of mercury, where the flasks were opened. (When the flasks are opened upside down in mercury the contents of the flasks doesn’t pour out, because mercury is much denser than water, and therefore the water will float atop of the mercury.)
Schwann used four glass tubes to inject air into the four opened flasks (one glass tube for each flask). Two of the glass tubes were at a red-hot temperature while air was injected through them. The two other tubes remained at room temperature during air injection. Schwann comments that An analysis had shown that air which had passed through a red-hot tube still contained 19.4 per cent oxygen. (For reference, normal air contains about 21% oxygen.) Therefore, such heating of the air does not significantly damage the oxygen in the air.
After the injection of air, the flasks were sealed and left alone. Schwann says that after 4 to 6 weeks, the liquid in the flasks which received unheated air had began to ferment. After twice as many weeks, there was still no sign of fermentation in the other flasks which received heated air.
However, when Schwann repeated this second experiment multiple times, he did not always get the same result. Pasteur quotes Schwann as saying that But in repeating these experiments, I found that they are not always so successful, and that sometimes fermentation does not occur in any of the flasks, for example, when they have been kept too long in boiling water, and sometimes, on the other hand, the liquid ferments in the flasks that have received heated air. (Schwann's inability to consistently replicate the results of this experiment will be explained later, by Pasteur.)
The results from Schwann’s two experiments showed that Gay-Lussac had been mistaken in his conclusion that the absence of [oxygen gas] is consequently a necessary condition for the preservation of animal and vegetable substances. Schwann showed that there is something in the air, something other than oxygen, which is necessary for the development of microorganisms in boiled liquids. And further, that this “something” is damaged by heating the air. But Schwann did not prove that this “something” was microbial germs. That last blow against the theory of spontaneous generation would be dealt by Pasteur.
Pasteur's experiments and the end of spontaneous generation
As was said previously, Pasteur's work on spontaneous generation was published in 1862, and an english Google-translation is AVAILABLE HERE. The text is divided into nine chapters, where the first chapter (“Historical background”) has already been referenced in previous sections.
Here are the two plates which accompany the text (the individual figures will be explained as we progress through the chapters):
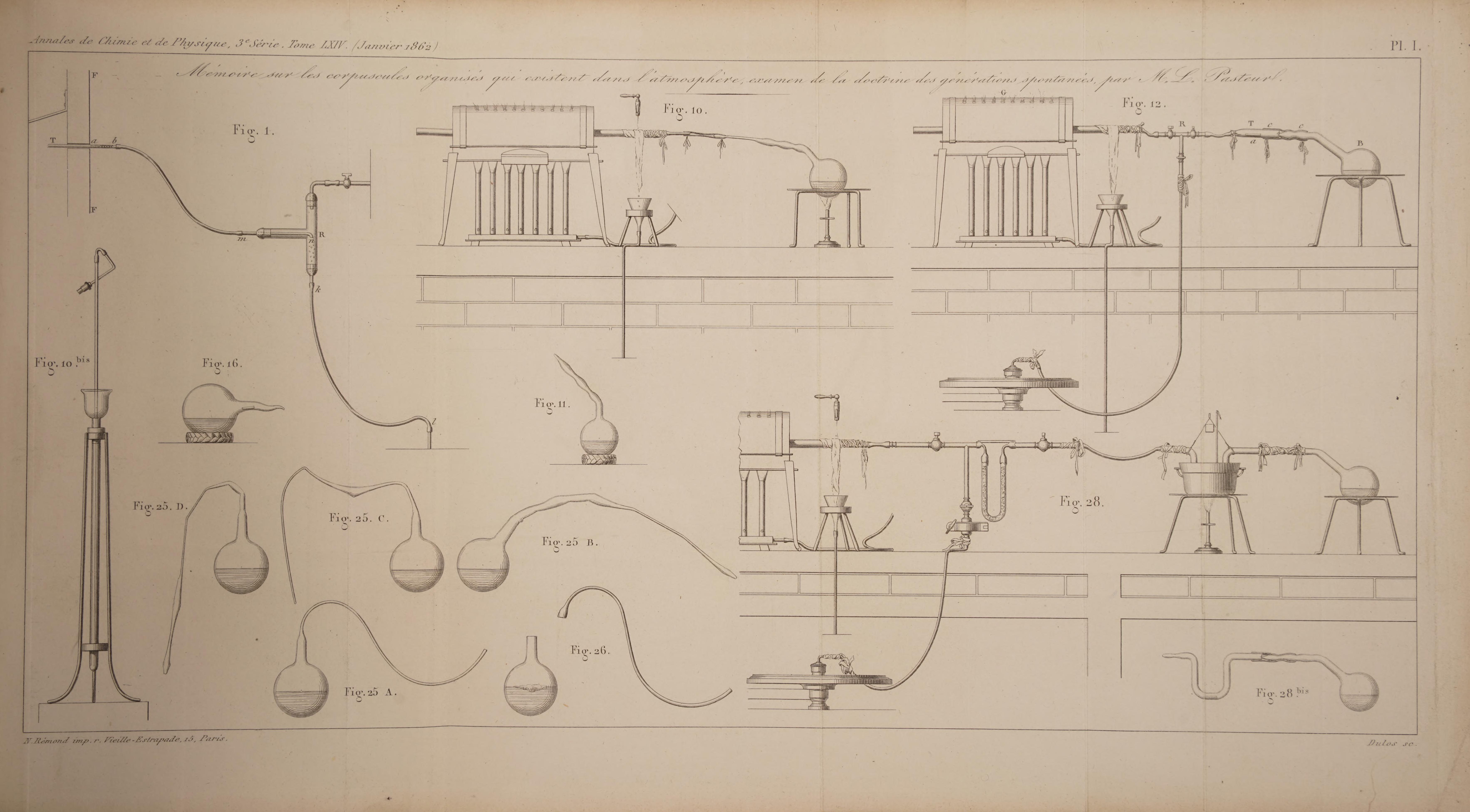
Source: Annales de chimie et de physique, 3rd series, volume 64, published 1862. Available on archive.org.

Source: Annales de chimie et de physique, 3rd series, volume 64, published 1862. Available on archive.org.
In his Chapter 2, Pasteur seeks to answer the following questions: Are there germs in the air? Are there enough of them to explain the appearance of organized growths [microorganisms] in infusions that have been previously heated? Can we get an approximate idea of the relationship between a given volume of ordinary air and the number of germs that this volume of air can contain?
He proceeds to describe a device he has constructed for sucking air into a dust-filter. (This device is shown in Pasteur's Plate I, Fig. 1, see the full translation previously mentioned for details on how this device works). As a filter he uses a piece of nitrocellulose, which has the important property that it will dissolve in a mixture of ether and alcohol. After trapping airborne dust particles in this filter, Pasteur dissolved the filter to release the collected dust. He then uses a microscope to look at drops of liquid containing these particles.
Various reagents can be applied to [the dust particles] using ordinary methods: iodine solution, potassium solution, sulfuric acid, and coloring agents. These very simple manipulations allow us to recognize that there is constantly a variable number of particles in the common air, whose shape and structure indicate that they are organized [“organized particles” basically means microorganisms]. Their dimensions range from the smallest diameters up to 1/100 to 1.5/100 and more millimeters [10 to 15 micrometer]. Some are perfectly spherical, others ovoid. Their outlines are more or less clearly defined. Many are completely translucent, but there are also opaque ones with internal granulations. Those that are translucent, with sharp outlines, so closely resemble the spores of the most common molds that the most skilled micrographer could not distinguish them.
That is all that can be said about them, just as one can only affirm that, among the others, some resemble encysted, ball-shaped infusoria, and generally the globules that are considered to be the eggs of these small organisms. But as for asserting that this is a spore, moreover the spore of a specific species, and that this is an egg, and the egg of a specific Microzoan, I believe that this is not possible. I confine myself, as far as I am concerned, to declaring that these corpuscles are evidently organized, resembling in every respect the germs of the lowest organisms, and so diverse in size and structured that they undoubtedly belong to a great many species.
A much more accurate idea of the number of corpuscles that their shape and volume allow us to call organized can be obtained by determining the average number of these corpuscles contained within the field of view of the microscope, and by knowing the ratio of the surface area of the droplet spread under the small glass slide covering it to the field of view of the microscope, for the magnification used. The total number of corpuscles in the droplet will be equal to the ratio we are discussing, multiplied by the average number of corpuscles contained within any given field of view. We can thus recognize that a small nitrocellulose wad exposed for twenty-four hours to the air current of the street, taken a few meters above the ground, during the summer, after a succession of fine days, gathers several thousand organized corpuscles for an intake of approximately one liter of air per minute.
Pasteur’s Plate II, Figures 2, 3 and 4, show “organized particles” in dust from the air, where the dust has been treated with different chemicals (iodine, potassium, etc.) in the different figures. Figs 6, 7, 8, and 9 shows various samples of dust, including both the organized particles and other dust particles.
In Chapter 3, Pasteur discusses the experiments of Schwann, in which a flask containing meat infusion or sugar water was boiled, before air was allowed to enter the flask through a heated tube. Schwann found that this prevented the emergence of microorganisms in the flask. Pasteur remarks that a number of scientists had attempted to replicate Schwann’s results, without success. Also:
I would add that Dr. Schwann himself was not always successful in his experiments on the inactivity of heated air; indeed, we saw in the first part of this Memoir, where I summarized this scientist's work, that his experiments on alcoholic fermentation often yielded results contrary to those he expected, without him being able to identify the presumed causes of error in these results.
Pasteur therefore repeats Schwann’s experiments. He takes a flask with a long thin neck, fills the flask partially with water containing sugar and “Albuminoid and mineral matter from brewer's yeast”. (The latter is called yeast extract, this is made by boiling and filtering yeast, then evaporating the water to leave a dry powder, the yeast extract). (Such a water solution, containing sugar and yeast extract, will be referred to as “sugar yeast water”). Sugar yeast water will normally give rise to microorganisms within a few days upon exposire to normal air. The neck of the flask is connected to a horizontally positioned tube of platinum, and this tube is surrounded by a heating apparatus. For an illustration of this setup, see Pasteur’s Plate I, Figure 10.
The platinum tube is heated red-hot, and the sugar yeast water in the flask is boiled for 2-3 minutes. Thus, all the air in the flask is either expelled by the steam, or is heated to 100°C by the steam. After the boiling stops, the steam in the flask condenses and the steam pressure is reduced, which allows outside air to enter the flask through the red-hot platinum tube.
When the flask has cooled down to room temperature, and is filled with air at normal atmospheric pressure, the flask is hermetically sealed by melting the glass neck with a flame. The flask is then placed in an area of temperature 30°C (which is favorable to the growth of microorganisms). Pasteur says that this method always succeeds in preventing the development of microorganisms in the sugar yeast water:
I affirm with the utmost sincerity that I have never had a single experiment, conducted as I have just described, that yielded a questionable result. Sugar yeast water, brought to a boil for two or three minutes and then exposed to air previously heated to red-hot temperature, does not deteriorate at all*, even after eighteen months at a temperature of 25° to 30°C. However, if left in ordinary air, after a day or two it is clearly deteriorating and becomes filled with bacteria, vibrios, or covered with mucors.
*) I have certainly had occasion to repeat this experiment more than fifty times, and in no case has this highly perishable liquid shown any trace of organized growth in the presence of heated air.
So why did other researchers, including Schwann himself, have trouble repeating Schwann’s experiment? Pasteur says their problem was their use of a mercury bath in their experiments. The surface of the mercury is exposed to the air in the laboratory and will consequently contain dust, including microbial germs. When, for example, a flask is submerged in the mercury bath (as in Schwann’s experiment), germs from the surface of the mercury will coat the submerged flask. And when the flask is opened, some of these germs will be also to enter into the flask.
(Note that Pasteur is able to prevent microbial growth in his flasks by boiling the liquids for just 2-3 minutes, whereas Spallanzani had to boil his infusions for nearly three quarters of an hour to prevent bacteria from appearing in his flasks. A possible explanation for this apparent disagreement could be that Pasteur's sugar yeast water is unsuitable for the growth of heat-resistant bacteria, so that even if some bacteria were able to survive the 2-3 minutes of boiling, they would not be able to grow. Alternatively, it could be that Spallanzani's beans and other vegetables contained heat-resistant bacterial spores, which were destroyed only when his vegetable infusions were boiled for at least 45 minutes.)
Recall Chapter 2, where Pasteur trapped dust particles in a nitrocellulose filter, and found that many of these particles resembled microorganisms. Now, if these dust particles truly are microorganisms, then microbial life should develop in sugar yeast water (and other nutrient liquids) that comes into contact with the dust. This is the topic of Chapter 4.
We know that when a liquid comes into contact with normal air, microbial life develops in the liquid, whereas if the liquid only contacts air which has previously been heated (“pre-heated air”), the liquid remains lifeless. The challenge is then: how to introduce dust particles into a liquid, without also introducing normal unheated air?
Pasteur’s experimental setup in this case is certainly clever (see his Plate I, figure 12). He uses one of the flasks produced by the method in Chapter 3 (where the flask is partially filled with sugar yeast water, boiled for a few minutes, and allowed to cool while heated air enters the flask, before the neck of the flask is sealed with a flame). The flask has been kept at 30°C for a couple of months with no sign of microbial life in the liquid (i.e. no clouding of the liquid, and no appearance of a microbial film or a clump of fungi).
A piece of nitrocellulose containing dust particles (labelled “a” in fig. 12) is placed in a glass tube (“T”), and this glass tube is connected to the neck of the flask (“B”) by means of a rubber tube. The glass tube containing the nitrocellulose is also connected to a branch of a T-tube (“R”), which has a valve at each of the three branches. The other two branches are connected to a vacuum pump and to a platinum tube with a heating apparatus.
In order to remove unheated air from the piece of nitrocellulose, Pasteur first closes the valve to the heated platinum tube, and opens the valve to the vacuum pump. Air is sucked out of the glass tube containing the nitrocellulose. Next, the valve to the vacuum pump is closed, and the valve to the heated platinum tube is opened. Heated air fills the glass tube containing the nitrocellulose.
Pasteur again closes the valve to the heated platinum tube and opens the valve to the vacuum pump, sucking the heated air out of the tube with the nitrocellulose. Next, the valve to the vacuum pump is closed again, and the valve to the heated platinum tube is opened again. The tube with nitrocellulose is again filled with heated air.
The vacuum and the re-entry of the heated air are repeated alternately ten to twelve times. The nitrocellulose is thus filled with heated air down to the smallest interstices of the fibers, but it retains its dust.
Once this replacement of unheated air for heated air is done, Pasteur breaks the neck of the flask containing the preserved sugar yeast water. The broken neck of the flask is still inside the rubber tube, and is therefore isolated from the outside air. The glass tube containing the piece of nitrocellulose is tilted sideways, causing the nitrocellulose to slide down into the flask through the broken neck. The neck is re-sealed with a flame, and the flask is stored at a temperature of about 30°C.
Each time Pasteur performs this experiment, signs of microscopic life appears in the flask in 24 to 48 hours. This is the same time required for such signs to appear in flasks exposed to normal air.
To verify that it is the dust particles, and not the dust filter itself, that causes development of life in the flasks, Pasteur replaces the nitrocellulose filter with an asbestos filter. He finds that when dust particles are trapped in the asbestos and the asbestos is placed in preserved sugar yeast water, then microbial life develops.
But with asbestos wads that had been previously heated and were not laden with dust, or with asbestos laden with dust but subsequently heated, no cloudiness, infusoria, or plants of any kind appeared. The liquids remained perfectly clear. I repeated these comparative experiments a great many times, and I was always surprised by their clarity and perfect consistency. It would seem, in fact, that experiments of this delicacy should sometimes offer contradictory results brought about by accidental causes of error. Yet, I have never once seen the blank experiments succeed, just as I have never seen the inoculation of dust fail to produce organized growth.
(I think nitrocellulose cannot be heated because it is explosive, which explains why Pasteur switches to using asbestos as a filter. It is not explained how he heats the asbestos and transfers it to the flask without contamination. But if we look at Pasteur’s Plate I, figs. 10 and 12, we can see that a part of the platinum tube, on the right side of the heating apparatus, is covered with a cloth with water flowing down upon it. It therefore seems that the heated air is cooled by this water before the air reaches the nitrocellulose or asbestos. I would guess that Pasteur heated the asbestos by simply turning off the water, thus letting un-cooled air reach the asbestos.)
In Chapter 5, Pasteur turns his attention from sugar yeast water to milk. In Chapter 2 he had found that if a flask partially filled with sugar yeast water is boiled for 2-3 minutes, allowed to cool while heated air enters the flask, and the neck of the flask is sealed with a flame, then this flask can be stored practically indefinitely without development of microorganisms. However, if the same procedure is performed on a flask of milk, then microorganisms will soon develop in the milk:
The experiments we have just described have always yielded similar results. Milk boiled at 100°C and left in contact with heated air fills, after a few days, with small infusoria, most often a variety of Vibrio lineola (Fig. 24, Plate II), and with bacteria, and, while retaining its alkalinity, it curdles.
I have never seen anything other than Vibrio and Bacterium form in milk treated in this way; no Mucedinaceae, no Torulaceae, no vegetable ferments [yeasts]. There is no doubt that this is due to the fact that the germs of these latter organisms cannot withstand 100°C in water, a fact I have also observed through direct experimentation. Similarly, we will recognize that if milk spoils under the preceding circumstances, it is because the infusoria we have just discussed can withstand the humid temperature of 100°C when the liquid in which they are heated possesses certain properties.
So a few minutes boiling at 100°C isn’t enough to prevent microbial growth in the milk. But what about a few minutes boiling at a temperature higher than 100°C? The boiling temperature can be raised by increasing the air pressure in the flask. Pasteur accomplishes this in the following way:
Let us return to the apparatus shown in Fig. 10, Plate I, and boil the milk at a temperature slightly above 100°C, 110°C at most, by attaching the glass tube from Fig. 10-bis [on the far-left side of Pasteur’s Plate I] to the left end of the platinum tube, immersing 40 to 50 cubic centimeters in the mercury of the long basin shown in that same figure. Let us detach this glass tube when the milk has boiled for only a minute or two; then let us close the neck of the flask with a flame, as we have always done.
(It seems to me that by allowing 50 cm3 of mercury to enter the glass tube (Fig. 10-bis), the air inside the closed system is compressed by the mercury, and this air compression causes an increase in air pressure. At least, I cannot think of any other explanation of how the mercury can increase the air pressure.)
When a flask of milk is prepared in this way, it can be kept indefinitely without any microbial growth. Pasteur also comments that boiling the milk at 100°C for a longer time can accomplish the same result.
But why is it that, compared to sugar yeast water, the milk must be boiled at a higher temperature or for a longer time in order to prevent the growth of bacteria? Pasteur, who had been educated as a chemist, considers that it may be due to acidity and alkalinity. Whereas milk is alkaline, sugar yeast water is slightly acidic.
And indeed, when sugar yeast water is mixed with some calcium carbonate, “which renders the liquid neutral or slightly alkaline”, Pasteur finds that bacteria can develop when the liquid has been boiled for a few minutes at 100°C. “If we now repeat these same tests, boiling the liquid at only 105°C, as we did earlier with the milk, we will see no cloudiness or mucorrhea whatsoever.” Pasteur thus considers it proved that an alkaline liquid gives the bacteria a slightly higher tolerance to heat, compared to an acidic liquid.
Further, if one takes milk (or alkaline sugar yeast water), which has been preserved by boiling at >100°C, and places a dust-loaded air filter into the milk (as was done in Chapter 4), then after a few days there will be visible signs of microbial growth in the milk. This shows that milk heated at >100°C is still fully capable of supporting microbial growth, and when milk heated at this temperature is preserved bacteria-free, it is not because the milk itself has been damaged in some way. rather, it is because the bacteria that were present in the milk were killed by the heat.
For anyone who is not yet fully convinced that microbial growth in preserved liquids is caused by germs in the air, in his Chapter 6 Pasteur has yet another experiment in store, and this one is simpler than any of the previous ones.
I place one of the following liquids, all highly perishable upon contact with ordinary air, into a glass flask: yeast water, sugar yeast water, urine, beetroot juice, or pepper water. Then, using a flame, I stretch the neck of the flask to create various curves, as shown in Fig. 25, Plate I.
I then bring the liquid to a boil for a few minutes until steam escapes abundantly from the open end of the tapered neck, without any other precautions. I then allow the flask to cool. A singular thing, quite likely to astonish anyone accustomed to the delicate nature of experiments concerning so-called spontaneous generation, is that the liquid in this flask will remain indefinitely unaltered.
No heating of the air that re-enters the flask is necessary, nor is it necessary to hermetically seal the flask. The long thin curved neck of the flask is all that is needed to prevent microbial growth in the liquid.
It seems that ordinary air, entering forcefully at first [once the boiling has stopped], must arrive in the flask completely untreated. This is true, but it encounters a liquid still close to boiling point. The air then enters more slowly, and when the liquid is sufficiently cooled to no longer deprive the germs of their vitality, the air entry slows enough that it leaves behind in the moist curves of the neck all the dust capable of affecting the infusions and causing organized growth. At least, I see no other possible explanation for these curious experiments.
If any such preserved flask has its long thin curved neck removed (see Pasteur’s Plate I, Fig. 26), then a development of microorganisms in the liquid will be apparent within a day or two, as is expected for boiled liquids exposed to normal air.
Here is a variation of the same experiment: liquid is placed into a flask, and the neck of the flask is stretched out as previously described. The liquid is boiled for a few minutes. While the liquid still boils, the end of the neck is sealed with a cork. The boiling is stopped, and the flask is allowed to cool down to room temperature. This causes the steam in the flask to condense to liquid water, so that a vacuum is generated inside the flask. Once the flask has fully cooled down, the cork is removed. The vacuum causes air to be sucked forcefully through the long thin curved neck, and microbial growth in the liquid then “usually appears after a few days”.
So, when air enters the flask slowly, the air stream through the long thin curved neck is not strong enough to carry germs all the way through the neck and into the liquid. Consequently, the liquid is preserved. But when air enters the flask forcefully, the air stream through the neck is strong enough to carry germs all the way through and into the liquid, and microorganisms therefore develop.
I must add that I currently have in my laboratory several highly perishable liquids, preserved for eighteen months in open vessels with curved and sloping necks, including several of those that were presented to the Academy of Sciences at its meeting of February 6, 1860, when I had the honor of communicating these new results to them.
The great advantage of this method is that it definitively proves that the origin of life in infusions brought to a boil is solely due to solid particles suspended in the air. Gases, various fluids, electricity, magnetism, ozone, known or unknown phenomena—there is absolutely nothing in ordinary atmospheric air, apart from its solid particles, that is the condition for the putrefaction or fermentation of the liquids we have studied.
At the beginning of his Chapter 7, Pasteur says that the experimenters at his time believed that even a tiniest bit of normal air was enough to cause the appearance of microorganisms in boiled liquids.
This view has always been supported, at least indirectly, by the practice adopted and deemed essential by observers of meticulously keeping ordinary air out of their experiments. As we have seen, sometimes they recommend heating the ordinary air, sometimes they subject it to energetic chemical agents; often they first place all its parts in contact with water vapor at 100°C (Spallanzani's experiment); finally, at other times they operate with artificial air [oxygen gas generated from water by means of electricity?], and, if it happens, under one of these various conditions, that the experiment gives rise to organized productions, they do not hesitate to assert that the operator has not been able to completely avoid the hidden influence of a small portion of ordinary air, however small it may be.”
Pasteur conducts a series of simple experiments which demonstrates the falsity of this belief. He takes a flask and partially fills it with sugar yeast water, which is then boiled for a few minutes. The neck of the flask is sealed with a flame, the boiling is stopped, and the flask is allowed to cool down. The condensation of steam creates a vacuum inside the flask. Pasteur then breaks off the neck of each flask, allowing a little bit of air to be sucked in, before he quickly re-seals the flask and places it in an oven with a temperature of 30°C. He repeats this process with a large number of flasks, with various types of liquids.
Here are the results of these experiments, which contradict generally accepted principles and, on the contrary, perfectly support the idea of germ dispersal. Most often, in just a few days, the liquid deteriorates, and the most varied organisms appear in the flasks, even though they are placed under identical conditions—far more varied indeed, especially with regard to Mucetaceae and Torulaceae, than if the liquids had been freely exposed to ordinary air.
But, on the other hand, it frequently happens, several times in each series of tests, that the liquid remains absolutely intact, regardless of the duration of its exposure in the oven, as if it had received heated air.
In this way he demonstrates that the component of air responsible for the appearance of microorganisms is not present in every small volume of air. That is, the responsible component is not spread continuously throughout the air. Rather, is it spread throughout the air as discrete particles, just like the germ-like objects which Pasteur found in airborne dust in Chapter 2. This is further evidence that the germ-like objects in the dust are in fact real germs.
Pasteur says that in these flasks which were only exposed to air for a short time, he finds a greater variety of microorganisms than in flasks which are exposed to air for a long time. This also makes sense, he thinks:
A small number of germs, in a limited volume of air, are not hindered in their development by more numerous germs or those with earlier fertility, capable of invading the space, leaving room only for themselves. Thus, Penicillium glaucum, whose spores are perennial and widespread, appears alone after very few days in unenclosed liquids, which, on the contrary, offer very diverse productions when subjected to limited quantities of air.
Additionally, Pasteur mentions that he opened and closed a series of ten flasks in a rarely used cellar, where the still air allows the airborne dust to settle onto the floor. The air in the cellar should therefore contain minimal dust. The result was that only one of the ten flasks gave rise to microorganisms, with the remaining flasks remaining lifeless.
On another occasion he opened and closed sets of twenty flasks in locations of varying altitude, including a place called Montanvert at an elevation of 2000 meters, and close to a glacier. Here also the result was that just one single flask gave rise to microorganisms.
These experiments should undoubtedly be multiplied considerably. But as they stand, they already tend to prove that as one ascends, the number of germs suspended in the air decreases noticeably. Above all, they demonstrate the purity, from our perspective, of the air on the high, ice-covered peaks, since only one of the vessels opened at Montanvert gave rise to a Mucédinea.
Assume that we know the temperature limit of fungal spores, the temperature where the spores lose their ability to develop into fungi. If airborne dust includes fungal spores, then dust subjected to heat below the spores’ temperature limit should retain its ability to cause fungal growth. On the other hand, dust subjected to heat above the spores’ temperature limit should lose its ability to cause fungal growth. To test whether this is indeed true is the topic of Pasteur’s Chapter 8.
For this experiment, Pasteur uses the setup illustrared in his Plate I, Figures 28 and 28-bis. A sealed flask partially filled with a preserved liquid is connected to a U-tube. A piece of asbestos (or cotton), which has been either brushed over the spores of a fungus or filled with airborne dust (by the method in Chapter 2), is placed into the U-tube.
The U-tube is connected to a T-tube with valves, which further connects to a vacuum pump and to a platinum tube with a heater. The T-tube also includes a sort of reservoir filled with concentrated sulphuric acid, which is used to dry the air inside the system. (Dry air at high temperature is less damaging to organisms than humid air at high temperature. Sulphuric acid binds strongly to water, which means that when water molecules in the air contacts the surface of the sulphuric acid, the water molecules will go into the acid and stay there, so that the air becomes dry. At least, this is my understanding of why Pasteur uses the acid and how it works.)
Pasteur uses the vacuum pump and the heated platinum tube to fill the system with heated air, similar to what he did in Chapter 4. The air inside the system should now be free from live germs.
Next, the U-tube containing the piece of asbestos is submerged into either boiling water (100°C), boiling salt water (108.4°C), or boiling oil (higher temperatures, I’m not sure what oil he uses, but he may use different oils as he achieves different temperatures in the range 119°C to 132°C). The heat from the boiling liquid is conducted through the dry air in the U-tube and to the asbestos. Thus, the asbestos is dry-heated at the temperature of boiling water or salt water or oil.
When the boiling is complete, the U-tube is removed from the boiling apparatus. The tip of the flask is broken with a hammer. (The flask is connected to the U-tube by means of a rubber tube, which doesn’t break when struck with the hammer, so the system remains sealed to the outside.) The U-tube is moved in such a way that the asbestos falls into the flask. (The U-tube connects to the T-tube by means of a rubber tube, so the U-tube can be moved without moving the rest of the system.) At last, the neck of the flask is re-sealed with a flame, and the flask is placed in an oven at a temperature of 20 to 30°C.
Pasteur gives the details of eight different experiments. Only experiments 4 to 8 are really of interest, but I will summarize all of them:
Experiment 1: Cotton with dust, dry-heated at 100° for 1 hour, then placed in preserved yeast water. Development of one kind of Mucorea and one kind of vibrio.
Experiment 2: Asbestos with dust, dry-heated at 100° for 15 minutes, then placed in preserved milk. Development of two kinds of infusoria.
Experiment 3: Asbestos with dust, dry-heated at 100°C for 1 hour, then placed in preserved sugar yeast water with chalk (calcium carbonate). Development of two kinds of vibrio.
Experiment 4: Asbestos with dust, dry-heated at 121°C for 30 minutes, then placed in preserved yeast water and in preserved urine. Development of fungi in both liquids.
Experiment 5: Asbestos with dust, dry-heated at 129°C for 30 minutes, then placed in preserved yeast water. No development of microbial organisms.
Experiment 6: Asbestos with Penicillium spores, dry-heated at 119 to 121°C for 30 minutes, then placed in preserved yeast water. Development of Penicillium (after more than 3 days).
Experiment 7: Asbestos with Penicillium spores, dry-heated at 108.4°C for 30 minutes, then placed in preserved yeast water. Development of Penicillium (after 2 days).
(Pasteur also notes that if the Penicillium spores are only heated at 100°C, or are not heated at all, then the spores germinate into fungi in just one day. Therefore, while dry-heating the spores at 108.4 or 121°C for 30 minutes doesn’t prevent them from developing into fungi, the high temperatures does affect the speed at which the spores can develop.)
Experiment 8: Asbestos with spores of Penicillium, and asbestos with spores of another fungus (Ascophora), dry-heated at 127 to 132°C for 30 minutes, then placed separately in two flasks of preserved yeast water. No development of microbial organisms in either flask.
In summary, I believe I can conclude from my experiments that the spores of common Mucedinaceae, heated in a vacuum or in dry air, remain fertile after being brought to a temperature of 120°C. It would probably be found that one could even go a little higher, perhaps to 125°C. Conversely, a fairly short exposure to 130°C is sufficient to render the spores of these same Mucedinaceae, which appear to be the most vigorous and the least sensitive, unfertile.
On the other hand, we find that the limits are the same for the fertility of airborne dust; that is, it produces Mucedinaceae even after being heated to 120°C, and it ceases to do so if subjected to a temperature of 130°C. The correlation of these results is further proof of the existence of Mucedinaceae spores among the organized particles that the microscope allows us to so easily identify in dust suspended in ordinary air.
This, then, is the end of our look at Pasteur's work on spontaneous generation. After Pasteur's long series of various experiments published in 1862, there was no longer any doubt: microorganisms which appeared in liquids were not generated “spontaneously” from the molecules in the liquids. Rather, the microorganisms originated with germs dispersed throughout the air. A century of uncertainty had come to a close.

Alcoholic fermentation
If water, sugar, and brewer’s yeast are mixed together, the sugar will disappear, to be replaced by alcohol (ethanol) and carbon dioxide (CO2). This conversion is called alcoholic fermentation. In the earlier part of the 1800s multiple scientists were seeking to understand the cause of alcoholic fermentation, and here we again find Louis Pasteur and his experiments to be of great significance. In fact, this entire topic will mostly be a summary of the second part of Pasteur’s 1860 article “Memoir on Alcoholic Fermentation” (“Memoire sur la fermentation alcoolique”). The french original is available on archive.org, and an English Google-translation is available here.
(Note: this topic, alcoholic fermentation, is perhaps more difficult to grasp than the other sections in this article. That's because this topic involves more chemistry, and I'm not much of a chemist, so I can't explain everything. Also, my reading on this topic hardly extends beyond that of Pasteur 1860. However, I still think this can be an interesting read.)
The first section of Pasteur's “Alcoholic Fermentation, Part 2” is a historical overview of fermentation. One of the first significant experiments on fermentation was done by a Mr. Thénard, who published his findings in 1803 (or possibly in 1802, I can’t tell which year it was, since it was published under the french revolutionary calendar). Pasteur writes:
By reducing the quantities in the single experiment reported by Mr. Thénard to 100 parts, 20 parts of fresh brewer's yeast and 100 parts of sugar, after complete fermentation, left 13.7 parts of an insoluble residue [yeast], still active, which, exhausted by contact with a new quantity of sugar, was reduced to 10 parts. This last residue [yeast] was white, exhibited all the properties of wood, and exerted no action on a new quantity of sugar water.
These results, to which I will return shortly, have been cited in all the works and have served as the basis for all discussions on the theory of fermentation. I must add immediately that there is, in Mr. Thénard's Memoir and even in the latest edition of his Treatise on Chemistry, a curious passage on the nitrogen in the ferment.
“New research,” he said, “worthy of the full attention of chemists, must be carried out on the decomposition that the ferment [yeast] undergoes. It will be necessary to see what becomes of the nitrogen in the decomposed ferment. It is not found mixed with carbon dioxide; it does not enter into the composition of the insoluble white matter; it is not part of a very small quantity of highly soluble matter found in the liquid with the alcohol. Alcohol does not contain any; so the question of what becomes of the nitrogen in the ferment is still to be resolved.”
Although this passage, as I shall show, contains several errors, it testifies to Mr. Thénard's preoccupation with the nitrogen in ferment, and moreover, to the fact that this eminent chemist, who had made a particular study of the subject, did not accept the opinions of authors on the transformation of nitrogen in yeast into ammonia [ammonia is a type of nitrogen-containing molecule]. It seems like it was Döbereiner who first announced that the nitrogen in yeast was present in the liquor in the form of ammonium salt, an assertion that was accepted by all chemists. It gradually made its way into elementary works, with the exception of Thénard's.
So, these are the main points from the extract above: if water, sugar, and yeast are mixed together and alcoholic fermentation proceeds until completion, then afterwards one finds that the yeast has lost weight during the fermentation. That is, it seems like the yeast has been partially decomposed. And the nitrogen-containing matter that was previously present in the yeast appears to have disappeared without a trace.
Some decades later, Charles Cagniard de Latour studied yeast under the microscope and found that the “globules” in the yeast appeared to grow and multiply. Thomas Brock, in his book “Milestones in microbiology”, translates some pages from Latour’s “Memoir on alcoholic fermentation” (“Mémoire sur la fermentation vineuse”, year 1838). Here is an extract:
I examined samples removed hourly from the fermentation vessel. One hour after the addition of the yeast, I already saw that the wort contained single globules each of which contained a small secondary globule attached to it. [Wort is a water extract of barley or other grain, containing sugar and other nutrients from the grain.] This small globule seemed to grow, since a little while later most of the pairs seemed to contain globules of the same size. The fourth sample revealed a very few paired globules. In order to assure myself that these pairs of globules were actually attached and not merely touching each other, I applied a force to the cover glass with a small needle while watching the globules under the microscope, and although this force produced a violent agitation of the globules, it did not affect the point of attachment of the pairs. It would appear that the individual globules become separated naturally as they grow to a certain age, since the commercial yeast generally consists of simple globules ...
(The development of new yeast cells is today called “budding”. Here is a video of budding.)
Latour says that, as “all brewers are aware”, when yeast is added to wort, the weight of the yeast increases during the fermentation. It was believed that this weight increase was due to plant proteins in the wort. During fermentation, the chemical changes taking place in the wort would reduce the solubility of the plant proteins. The proteins would then precipitate into a solid mass, which became mixed with the solid mass of yeast, thus adding to the apparent weight of the yeast. Further:
It has been reported to me that it is possible to cause the production of globules when egg white is diluted in water and heated, and that the globules of yeast arise from a nitrogenous material of animal or vegetable origin, which is present in the wort, and coagulates to form the globules. If this is so, the yeast globules are no more living than those globules obtained during the coagulation of egg white by heat.
Based on his own microscopic examination, he has found that while the globules in the heated egg white has about the same size as the globules in yeast, they do not look entirely similar. In particular, the globules in the egg white lack the internal granules which can be seen in the globules of yeast.
It seems to me, therefore, that the objection that the globules of ferment [yeast] are analogous to those of coagulated egg white is not valid.
Based on this observation, and on the apparent growth and multiplication of the yeast globules as seen under the microscope, Latour thinks that the increased mass of yeast during fermentation is caused not by coagulation of plant proteins, but by the growth and multiplication of the yeast globules. Consequently, he considers the yeast globules to be living organisms. He also believes that the decomposition of sugar and appearance of alcohol and carbon dioxide are effects of the growth of the yeast globules, where the sugar is used as a nutrient. However, he cannot prove this latter hypothesis.
According to Pasteur, Latour’s idea that yeast is a living organism that feeds on sugar and produce alcohol and carbon dioxide as metabolic products, “immediately found a powerful opponent in Mr. Liebig”. Liebig was a chemist, and believed that fermentation was the result not of a process of life, but of a process of decomposition. Basically, he believed that when a substance of animal or plant origin is decomposing, and this decomposing substance is in contact with sugar, the state of decomposition can spread to the sugar, causing the sugar to split into alcohol and carbon dioxide. By 1860, Liebig’s ideas on fermentation were “generally accepted in Germany and France”.
Pasteur continues:
... In my opinion, this is the main reason for the gradual success that Mr. Liebig's ideas have acquired among chemists. For twenty years, a great number of phenomena have been discovered that are classified alongside alcoholic fermentation proper, and in which it has seemed impossible to recognize the existence of particular cryptogamic plants [yeasts], but in all of them there was matter, once alive, in the process of decomposition.
(Note: yeast are not classified as plants in modern biology.)
And, for example, if one places dissolved sugar mixed with chalk with any nitrogenous animal matter—caseum, fibrin, gelatin, rennet, an animal membrane, etc.—one sees the sugar gradually become lactic acid. Now, these animal substances are of very diverse structure, nature, and form, and the final effect on sugar is the same. Only one thing appears to be similar in these nitrogenous substances: their gradual decomposition. The correlation is thus evident between the transformation of sugar into lactic acid and a process of decomposition of a nitrogenous substance.
Research by Mr. Colin on alcoholic fermentation, dating from 1825, had already established similar facts. This chemist had recognized that a wide variety of animal substances could cause sugar to split into alcohol and carbonic acid.
(Carbonic acid is the water-dissolved form of carbon dioxide.)
However, a remarkable circumstance should have aroused attention and prompted caution, at least concerning alcoholic fermentation. Indeed, after the publication of Mr. Cagniard de Latour's observations, Mr. Turpin, who had been tasked with reporting them to the Academy of Sciences, studied, at Mr. Thenard's request, the deposit that forms during the alcoholic fermentation of sugar by decoction of egg white, and found that it consisted solely of brewer's yeast globules. If one of the substances used by Mr. Colin, egg white, only caused alcoholic fermentation by producing yeast, it could be presumed that all other nitrogenous substances behaved similarly, and therefore their diversity proved nothing regarding Mr. Liebig's theory.
But I hasten to add that nothing of the sort existed, or so it was believed, in the very diverse and numerous cases of lactic fermentation. All observers agree that there is only a chemical decomposition of the animal matter. The facts relating to this fermentation therefore had a decisive influence on the theory.
Thus, the idea of Mr. Cagniard de Latour, which had initially enjoyed some credibility, was gradually abandoned. Many people, at least, did not dispute that brewer's yeast was organized [living], but they believed that it was partially decomposed by fermentation, as Mr. Thénard had stated, and that, resembling all other nitrogenous substances acting as fermenting agents, it was to this property that it owed its action on sugar. This was Mr. Liebig's view.
“If fermentation,” says Mr. Liebig, “were a consequence of the development and multiplication of the globules, they would not excite fermentation in pure sugar water, which lacks the essential conditions for the manifestation of vital activity. This water does not contain the nitrogenous matter necessary for the production of the nitrogenous part of the globules. The globules cause fermentation, not because they continue to develop, but as a result of their nitrogenous internal part which decomposes into ammonia and other products, that is to say, as a result of a chemical decomposition, which is the complete opposite of an organic process.”
So, this was the state of the science when Pasteur began his work on alcoholic fermentation.
In Thénard’s experiment on fermentation in sugar water, Thénard had not been able to determine what had happened to the nitrogenous matter in the yeast during fermentation. After a completed fermentation, he was not able to detect any nitrogen in the solids (exhausted yeast), nor in the fermentation liquid, nor in the fermentation gas (carbon dioxide).
Another scientist, Döbereiner, believed he had found that the nitrogenous matter from the yeast was present in the fermentation liquid in the form of ammonum salts. By treating the fermentation liquid with “alkalis”, the ammonium salts could be converted to free ammonia, and this ammonia could then be quantified.
(According to the Encyclopedia Britannica article on alkali, an “alkali” is any of the soluble hydroxides of the alkali metals—i.e., lithium, sodium, potassium, rubidium, and cesium. Alkalies are strong bases that turn litmus paper from red to blue; they react with acids to yield neutral salts; and they are caustic [causing chemical burns] and in concentrated form are corrosive to organic tissues. The term alkali is also applied to the soluble hydroxides of such alkaline-earth metals as calcium, strontium, and barium and also to ammonium hydroxide. The term was originally applied to the ashes of burned sodium- or potassium-bearing plants, from which the oxides of sodium and potassium could be leached.)
Pasteur's recount is a little unclear on the following point, but as I interpret it, Döbereiner quantified the ammonia in the liquid before and after fermentation, and found that the ammonia content had risen during the fermentation. This then supported the hypothesis that the yeast is partially decomposed during fermentation.
Regarding Döbereiner's results, Pasteur has the following to say:
Döbereiner's results are erroneous. The soluble residue of the fermented liquid does not contain the slightest amount of ammonium originating from the nitrogenous matter of the yeast. The nitrogen that the fermentation liquid contains is entirely in the form of nitrogenous organic matter, highly susceptible to alteration by alkalis, and even releasing ammonia when exposed to potash, lime, and baryte at room temperature. This is what misled Döbereiner.
My own experiments on the nitrogen in ferments only achieved the desired precision once I became aware of the excellent methods for determining ammonia described by Mr. Boussingault in his Memoirs and in his remarkable lectures at the Conservatoire des Arts et Métiers, where I was able to witness every detail of the procedures. Mr. Boussingault's most valuable information for the research I had undertaken is the following: calcined magnesium converts ammonium salts to free ammonia at boiling point, but does not release ammonia from nitrogenous organic matter susceptible to alteration by lime, potash or baryte.
... That being said, here are some analyses of the fermentation liquids. They clearly show that not the slightest amount of ammonium is formed during the action of the yeast [...] 10 grams of sugar were fermented with 200 cubic centimeters of clear yeast water containing 0.0075 grams of ammonia and a trace of fresh yeast. Six days later, by testing the entire liquid, it was found to contain 0.0005 grams of ammonia. The consistency of the results of these experiments and many similar ones leaves no doubt as to the main fact. Not the slightest amount of ammonia is formed during alcoholic fermentation at the expense of the yeast. But these results go further: they indicate the disappearance of a portion of the ammonia present in the initial liquid. This latter circumstance led me to add ammonia directly in order to better study the phenomenon.
The following experiments were carried out in this direction: 100 grams of sugar was fermented with 10 grams of washed yeast paste and 0.200 grams of ammonium L-tartrate, containing 0.0185 grams of ammonia. The fermentation lasts a very long time. When all the sugar [...] has disappeared, the liquid is analyzed. It contains only 0.0015 g of ammonia. I also found free tartaric acid in the solution with all its characteristics. Thus, almost all the ammonia added in the form of ammonium tartrate has disappeared, as has that which was present in the 10 grams of yeast.
Based on this result, Pasteur wonders if the ammonium disappears because the yeast combines ammonium with carbon from sugar, so as to create new nitrogenous organic matter (which the yeast needs to grow). To test this hypothesis, he creates a solution composed of sugar, ammonium tartrate, and yeast ash (which contains minerals also needed for growth). This solution does not contain any nitrogenous organic matter, unlike the solutions of yeast water he used in previous experiments. And he finds that if a tiny amount of fresh yeast is added to such a solution, the yeast is able to multiply:
... Over the following days, the liquid gradually becomes cloudier, as does the release of gas, which becomes noticeable enough for foam to fill the neck of the bottle. A deposit gradually covers the bottom of the container. Observed under a microscope, a drop of this deposit reveals a beautiful, highly branched yeast, extremely young in appearance; that is to say, the globules are swollen, translucent, and not granular, and one can distinguish, with surprising ease, each globule from the small quantity of yeast sown at the beginning. These globules have a thick shell, standing out in darker circles; their contents are yellowish and entirely granular; but the way in which they are sometimes surrounded by the young globules clearly indicates that they have given rise to those of the latter that form the heads of the strands.
It is in the first few days that these interesting observations must be made: in the evening, by the bright light of the gaslight, the old globules stand out from the infinitely more numerous young ones, just as one would distinguish a black marble among many white ones. Gradually, the differences disappear, and the newly separated globules lose all branched appearance; buds are no longer visible. The globules are very granular, like mature or exhausted brewer's yeast.
Moving on, Pasteur says that when fresh yeast is transferred to pure water, and subsequently filtered out of this water, an analysis of the water shows that it is no longer pure. Rather, the water now contains nitrogenous organic matter as well as minerals. This nitrogen and minerals could only have come from the yeast, meaning that fresh yeast can carry these substances within itself.
Therefore, when fresh yeast is transferred to pure sugar water, the yeast not only has access to sugar, but also to nitrogenous organic matter and minerals. That's all the yeast needs to grow and multiply, as demonstrated by Pasteur's previous experiments.
Pasteur now quotes Liebig on fermentation, and while I gave this quote in the section on the history of fermentation research, it should be added here again:
“If fermentation,” says Mr. Liebig, “were a consequence of the development and multiplication of the globules, they would not excite fermentation in pure sugar water, which lacks the essential conditions for the manifestation of vital activity. This water does not contain the nitrogenous matter necessary for the production of the nitrogenous part of the globules. The globules cause fermentation, not because they continue to develop, but as a result of their nitrogenous internal part which decomposes into ammonia and other products, that is to say, as a result of a chemical decomposition, which is the complete opposite of an organic process.”
Pasteur’s experiments shows that Liebig was mistaken. When yeast is present in a liquid containing all the nutrients required for its growth (sugar, nitrogen-containing matter, and minerals), the yeast can ferment sugar and multiply itself for as long as all three nutrients are present. If this yeast is filtered out of this nutrient-rich liquid and transferred to pure sugar water, the yeast carries with it some nitrogen-containing matter and some minerals. The yeast can therefore continue to ferment sugar and multiply itself in this liquid that did not originally contain any other nutrient than sugar. However, the nitrogen and minerals which the yeast carried with it will eventually be completely consumed, and the yeast is then no longer able to ferment the remaining sugar in the liquid, and it no longer multiplies.
Let us now return to Thénard’s experiment, mentioned in the beginning of this section. Thénard’s found that 20 parts of brewer's yeast and 100 parts of sugar, after complete fermentation, left 13.0 parts of an insoluble residue [old yeast] which, when brought into contact with a new quantity of sugar, was reduced to 10 parts. Many people believed this weight loss was evidence that the yeast was decomposing, and that the decomposition was causing the fermentation.
But Pasteur has found that when yeast is filtered out of a nutrient-rich liquid and moved to sugar water, a part of the nitrogen-containing matter and minerals which the yeast carries with it will “leak out” of the yeast and into the surrounding sugar water. It is the loss of this soluble nitrogen and minerals which causes the apparent weight loss of the yeast. If one weight the yeast before and after fermentation in sugar water, and then one also determines the weight of the dissolved nitrogen-containing matter in the liquid after fermentation, one finds that the “total yeast mass” (yeast after fermentation + dissolved nitrogen) has increased, not decreased.
Pasteur determines that the increase in total yeast mass is equal to roughly 1.2 to 1.5% the mass of fermented sugar, so that if 100 grams of sugar is fermented, the total yeast mass increases by 1.2 to 1.5 grams. (The yeast uses most of the sugar for energy, while a small part of the sugar is converted into “building material” for the yeast cells, and it is this added building material that causes the increase in total yeast mass.)
When fermentation occurs in a liquid containing sugar as well as a good amount of nitrogenous organic matter and minerals, then the total yeast mass also increases. But in this case, the increase in total yeast mass is equal to about 1% of the mass of fermented sugar, slightly lower than for a fermentation in pure sugar water. (Pasteur doesn’t know why the increase in total yeast mass differs slightly between pure sugar water and sugar water with nitrogen and minerals.)
Pasteur therefore showed what Charles Cagniard de Latour had hypothesized when he discovered that yeast is a living organism: that alcoholic fermentation, the conversion of sugar into alcohol and carbon dioxide, is a result of the growth of the yeast globules, where the yeast uses the sugar as a nutrient.
Bacteria as a cause of disease, and the discovery of solid culture methods
On the topic of bacterial infectious disease, the major historical figure is Robert Koch. Koch is most famous for having proved that tuberculosis, one of the most lethal diseases of the past, is caused by a specific species of bacteria which Koch called the “Tubercle bacilli” (today they are known as “Mycobacterium tuberculosis”).
To learn about the history of disease-causing bacteria, we can start with a lecture given by Koch in 1890, with the title “On Bacteriological Research: Lecture at the 1st General Session of the 10th International Medical Congress on August 4, 1890” (“Ueber bakteriologische Forschung: Vortrag in der 1. allgemeinen Sitzung des X. internationalen medicinischen Congresses am 4. August 1890“). The original German text is available on archive.org, while an english Google-translation is available here.
This is Koch’s opening of the lecture:
Bacteriology, at least as far as it is relevant to us physicians, is a very young science. Just fifteen years ago, little more was known than that peculiar, foreign-like structures appear in the blood in cases of anthrax and relapsing fever, and that so-called vibrios [a type of bacteria] occasionally occur in wound infections. No proof had yet been provided that these substances could be the cause of those diseases, and with the exception of a few researchers, such findings were regarded more as curiosities than as potential pathogens. This was hardly surprising, since it had not even been proven that these were independent organisms specific to these diseases. Bacteria indistinguishable from anthrax bacilli had been found in putrefying fluids, particularly in the blood of suffocated animals. Some researchers refused to consider them living organisms at all, instead viewing them as crystalline structures. Bacteria identical to relapsing fever spirochetes were said to be found in swamp water and dental mucosa, and bacteria identical to the micrococci of wound infection diseases were allegedly found in healthy blood and healthy tissues.
As Koch explains, progress first began when new microscope technology and new dyes for coloring bacteria became available in the 1870s. Bacteria in water had been seen since Leewenhoek in the late 1600s, but these new tools of the 1870s were needed to properly study bacteria located inside animal tissues.
(If you read my article on the history of cell biology, you may remember that the ability of scientists to study chromosomes was improved in the 1870s due to the introduction of new microscope technology and dyes. So these same tools led to simultaneous progress in the study of bacteria and in the study of chromosomes. However, it is possible that different dyes were used to color bacteria on the one hand and chromosomes on the other hand, I am not sure about this.)
Koch continues:
The idea that microorganisms must be the cause of infectious diseases was indeed expressed very early on by some outstanding minds, but the general public was not yet comfortable with it and remained very skeptical of the first discoveries in this field. All the more reason, therefore, to provide irrefutable proof, especially in the early cases, that the microorganisms found in an infectious disease were indeed the cause of that disease. At that time, the objection was still valid that it could be a matter of chance encounter between disease and microorganisms, that the latter played the role not of dangerous parasites, but of harmless parasites which were unable to live in a healthy body, and which found the right conditions for life only in diseased organs. Some acknowledged the pathogenic properties of bacteria, but considered it possible that they initially were harmless organisms which had only transformed into pathogenic bacteria under the influence of the disease process.
If, however, it could be demonstrated: firstly, that the parasite is found in every single case of the disease in question, and indeed under conditions that correspond to the pathological changes and the clinical course of the disease; secondly, that it occurs in no other disease as an accidental and non-pathogenic parasite; and thirdly, that, completely isolated from the body and sufficiently often cultivated in pure cultures, the parasite is capable of reproducing the disease; then it could no longer be an accidental side effect of the disease, but in this case no other relationship between parasite and disease could be conceived than that the parasite is the cause of the disease.
Regarding the third point: in infectious diseases like tuberculosis, people knew that the disease could be transmitted between animals by taking a piece of sick tissue from one animal and inserting it into a wound on a healthy animal. If a certain species of bacteria was found to be present in the sick tissue, then one could hypothesize that this species of bacteria was the cause of the infection. But such infection experiments did not constitute proof that the bacteria were responsible for infection, because the sick tissue contained other things apart from the bacteria. One may consider that some sub-microscopic particle could be the real cause of the disease.
To substantiate the hypothesis that the bacteria were the cause of illness, it was necessary to first obtain the suspected bacteria free from any accompanying sick tissue. That is, it was necessary to grow the bacteria in a laboratory, to obtain a “laboratory culture” of the bacteria. If such a laboratory culture, after being transferred to test animals, would cause these animals to develop the disease, then this was much better evidence that these bacteria were the real cause of the infection.
Notice how Koch says that the bacteria should be cultivated in “pure cultures”. A pure culture is a culture that only contains a single species of microorganism, for example, a culture that only contains the bacteria from tuberculous tissue, free from other microorganisms. If one of the many microorganisms which are present in the air, on the surface of objects, or in liquids, ends up in a culture and starts growing there, then the culture is contaminated (i.e. no longer pure).
If you were to perform infection experiments with a bacterial culture and your test animals became sick, but you were using contaminated cultures to infect the animals, then you can’t really say that “this particular bacteria is the cause of the disease”. Any of the contaminating microorganisms could be involved in the process.
When Koch began his work on bacteria, all bacteria were grown in liquid cultures, and there existed no good method for keeping a liquid culture pure. Koch discovered that if the bacteria were cultured on a solid surface, such as on a potato slice, then it was much easier to obtain and maintain purity. The details of his work with solid cultures were presented in his 1881 article “For the investigation of pathogenic organisms” (“Zur Untersuchung von pathogenen Organismen”). The original German article is available on archive.org, while an english Google-translation is available here.
In that article, in the section on “Pure culture”, Koch begins by by describing the many ways in which a liquid culture can become contaminated, and the methods some scientists have employed to try to cope with the issue.
It is quite obvious that all efforts in this direction are futile. Therefore, I have completely abandoned the principle of pure culture I have followed until now and embarked on a new path. A simple observation, which anyone can easily repeat, led me to this point.
If you halve a cooked potato and leave it cut-side up in the air for a few hours, then place it in a humid room, for example under a moistened glass bell jar to prevent it from drying out, you will notice, depending on the temperature of the room, on the following, second, or third day, that various very small droplets form on the potato, almost all of which seem to be different from one another. Some of these droplets are whitish, porcelain-like in color; others are yellowish, brown, light gray, or reddish; some look like flattened water droplets; others are hemispherical; and still others are wart-like. But all of them enlarge to a greater or lesser extent, and mycelium of mold spores appears among them. Finally, the individual droplets merge, and soon the potato begins to rot.
If these droplets are now examined microscopically while still isolated, ideally after being spread on a coverslip, heated, and stained, it turns out that each one consists of a specific type of microorganism. For example, one droplet contains large micrococci, another very small ones, a third micrococci arranged in chains, while other colonies, especially the flat, membrane-like ones, are formed by bacilli of varying sizes and arrangements. Some consist of yeast cells, and occasionally, the mycelium of a mold sprouting from a spore is found.
The origin of all these different organisms will not be in doubt for long if another potato, peeled with a hot knife to remove the bacilli spores that always adhere to the peel along with the soil and are not killed by the brief boiling, is kept away from the air in a disinfected beaker with a cotton wool plug and observed. No droplets form on the potato treated in this way, no organisms settle on it, and it remains unchanged until it gradually dries out after several weeks. Therefore, the germs that developed into the small, droplet-like colonies on the first potato could only have settled from the air, and one often finds, indeed, a clearly recognizable dust particle or fiber in the center of the small colony, which served as a carrier for the germs, be they dried, still viable bacteria, yeast cells, or spores.
However, to avoid any misunderstanding, I must add that on the unpeeled potato, individual colonies develop from the edges, arising from germs that are located on the peel, specifically in the soil adhering to it.
What conclusions can be drawn from this observation of the colonies growing on the potato? Most striking is the fact that, with a few exceptions—where presumably two different germs were so close together that the resulting colonies were bound to merge, or where the same speck of dust was laden with different germs and these began to develop simultaneously—each droplet or colony is a pure culture and remains so until, during further growth, it collides with its neighbor and the individuals of one colony mix with those of another.
If, instead of the potato, the same surface area of a nutrient solution had been exposed to the air, then germs from the air would undoubtedly have settled on its surface, and indeed approximately the same number and the same types as on the potato. The development of these germs would have proceeded in a completely different manner than previously described. The motile bacteria would have rapidly dispersed in the liquid, mingling with the initially somewhat cohesive, floating colonies of immotile bacteria. Their lively movements would have also disturbed and scattered these colonies far and wide. Some organisms would drift at the bottom of the liquid, others in the upper layers. Some that found a spot on the potato where they could multiply undisturbed would be suffocated by the more vigorously growing organisms in the nutrient solution as soon as they began to grow, and would never develop at all.
In short, from the very beginning, a microscopic examination of the entire liquid would reveal a chaotic mixture of forms and shapes, never even remotely resembling a pure culture. But what is the fundamental difference between the nutrient medium that the potato offers the microorganisms and that provided by the nutrient solution? The difference lies solely in the fact that one is solid and prevents the different species, even if they are mobile, from being mixed together, while in the other liquid nutrient substrate, there is no question of the species remaining separate at all.
Now, if one wishes to obtain pure cultures of bacteria from sick tissue or from blood, then one would take some tissue on the point of a needle and streak this across the surface of a sliced boiled potato. Except, as Koch finds, many bacteria obtained from animal tissue don’t grow on potato. This problem he solved by taking a liquid nutrient solution, in which bacteria from animal tissues can grow, and adding gelatin to create a solid “nutrient gelatin”. For example, blood serum can be mixed with gelatin to create “blood serum gelatin”. A piece of animal tissue containing bacteria can then be streaked across the surface of the blood serum gelatin. Bacterial colonies will develop on the solid surface of the serum.
While different species of bacteria can be differentiated based on the size, color and shape of their colonies, it is also possible to use a colony’s microscopic appearance to determine the species of its bacteria.
For example, anthrax bacilli and hay bacilli in gelatin cultures are not at all confusable. The anthrax bacilli are never motile and always form clumps consisting of long, wavy, and curly threads, often twisted around each other. Hay bacillus, on the other hand, only develop into longer filaments in very young colonies. As soon as they develop further and, as is regularly the case, liquefy the gelatin, they are seen only as lively, mobile rods filling the interior of the colony and, at its periphery, burrowing into the still-solid gelatin in perfectly regular masses directed perpendicularly to the periphery, so that the colony appears as if surrounded by a radiating halo.
This creates such a characteristic image, so different from that of the anthrax colony, that one can immediately recognize both types of bacillus among all other microorganisms by the described characteristics. Other bacillus exhibit yet other forms; the mobile ones mostly form wreath-like figures similar to those of the hay bacillus, but differing from it in the shape and width of the radiating halo. Still other bacillus form colonies that resemble a far-reaching, intricately intertwined root system. In the disinfection experiments, we obtained one of the Bacillus strains that withstood heat the longest. It is rather plump and forms flat colonies on the gelatin, in which the individual bacilli are arranged close together in a mosaic pattern, thus forming no pseudo-filaments and showing no movement.
Later, when Koch was studying tuberculosis, the above-described method of pure culture was used to infect test animals with pure Tubercle bacilli, and in this way Koch could demonstrate that the “Tubercle bacilli” was the actual cause of tuberculosis. (The actual details of Koch's work on Tuberculosis aren't really that interesting, at least not to me, so I choose not to include those details here.)
(For those interested in having a look, Koch’s work on tuberculosis was published in 1884 under the title “The etiology of Tuberculosis” (“Die Aetiologie der Tuberkulose”), available in the German original on Google books (see pages 1 to 88). An 1890 English translation by a Mr. Saure is available on archive.org.)
To this day, people who work with microorganisms are still using nutrient gel cultures in the manner described by Koch. However, gelatin has been replaced by another gelling agent called agar. Gelatin gels become liquid in certain circumstances, and I believe agar gel is preferred because it is less prone to liquefying.
Summary
At long last this article comes to an end, and with it we've covered quite a good bit of history. On the topic of spontaneous generation there was Needham's experiments with sealed, heated infusions, whose results led him to believe that particles of organic matter contain a “vital force”, which allows these particles to assemble into tiny organisms. Similar experiments by Spallanzani found that hermetically sealed vegetable infusions must boil for three quarters of an hour to prevent the growth of microorganisms.
Gay-Lussac analyzed the air in hermetically preserved food and found that this air contained no oxygen. He therefore believed that a lack of oxygen was a prerequisite for preventing microbial growth in canned food. Schwann performed experiments in which heated air, still rich in oxygen, was passed into vessels with meat infusion and sugar water, with the result that no microbial growth took place. This refuted Gay-Lussac's conclusion on the necessary absence of oxygen in canned food.
At last, there was Pasteur, who filtered air through nitrocellulose, then dissolved the filter and looked at the dust particles through his microscope. Many of the dust particles were visually indistinguishable from microorganisms. He then performed a number of experiments to prove that these dust particles were in fact microorganisms. His results showed that in order to prevent microbial growth in a liquid, it is both necessary and sufficient to: 1. Destroy the germs already present in the liquid, which is done by boiling, and 2. Prevent airborne germs from entering the liquid.
The next topic was the role of yeast in alcoholic fermentation. One influential early experiment on this matter was conducted by Thénard, who found that when fresh yeast is moved to pure sugar water, fermentation will begin, but after a while the yeast loses its ability to cause fermentation. In addition, it appeared that the yeast was losing weight during the fermentation.
Cagniard de Latour was the first person who witnessed the process of yeast budding, where new yeast globules are created from the old. He concluded that the yeast was a living organism, and hypothesized that fermentation was a consequence of the yeast using sugar as a nutrient.
At around the same time, Liebig began to champion the idea that fermentation is caused by decomposition, where a state of decomposition is transferred from one substance over to sugar. In the case of yeast, he thought that it was the decomposition of the nitrogenous organic matter in the yeast that led to the decomposition of sugar.
Pasteur found that the apparent weight loss of the yeast in Thénard's experiment was caused by the nitrogenous organic matter of the yeast leaking out into the water. When the weight of this nitrogen was accounted for, the yeast was found to increase in weight during fermentation, equal to about 1 to 1.5% the weight of fermented sugar. Pasteur's work thus supported Cagniard de Latour's idea that the yeast is consuming the sugar.
The final topic was infectious disease and solid cultures. According to Koch, work on the connection between certain bacteria and certain infectious diseases began in the 1870s. A number of people believed that bacteria may not be the true cause of disease, even if they were found to be associated with diseased tissues.
To respond to such scepticism, and to prove that a certain bacteria truly is the cause of a certain disease, Koch thought it necessary to obtain pure laboratory cultures of the given bacteria, and to show that injection of such pure bacteria into test animals led to the animals developing the disease.
Obtaining and maintaining pure cultures in nutrient liquids like blood serum was very difficult. Koch first discovered that a sliced boiled potato, exposed to air, would permit bacteria to grow as isolated colonies, where each colony essentially was a pure culture of a single bacteria. He further found that nutrient liquids like blood serum could be used to create solid cultures simply by adding gelatin. With the aid of the solid culture method, Pasteur was able to create compelling evidence that the bacteria he called the Tubercle bacilli were the true cause of tuberculosis.